

The DAWNDINOS team, spearheaded by Dr. Peter Bishop, have now published two papers on the locomotion of extinct archosaurs, in the journal Paleobiology. These studies advance our project towards its major goals of reconstructing how locomotion evolved in Archosauria, such as the multiple origins of bipedalism, and advancing methods in biomechanical modelling and simulation in order to better test whether Triassic dinosaurs had some form of “locomotor superiority”, such as faster running speeds, relative to pseudosuchian archosaurs.
The first paper used 3D digital data on body dimensions (Figure 1: body mass, fore- and hindlimb length, and centre of mass position) of 80 archosaurs and close relatives to estimate, via a battery of statistical morphometric tests, whether certain archosaurs were bipedal or quadrupedal. Importantly, we applied this method to species that were known bipeds/quadrupeds and obtained good precision in doing so; boosting confidence in our method.

We found (Figure 2) that our study species Batrachotomus was quadrupedal (as expected), whereas Postosuchus (a somewhat controversial animal) and Marasuchus (as expected) were bipedal. Interestingly, Euparkeria was in an unusual area of the “morphospace” that was ambiguous but closer to the quadrupedal domain. Overall, our methods tended to support our parallel study indicating that the dinosaur Mussaurus transitioned from a quadrupedal hatchling to a bipedal adult. This analysis also raised fascinating unexpected questions because the juvenile Alligator was consistently misclassified as a biped, which we concluded was due to its longer limbs and lighter head. The variations in body shape, including limb bone dimensions used in analogous studies, across growth might thus be giving scientists misleading signals about locomotor ability.
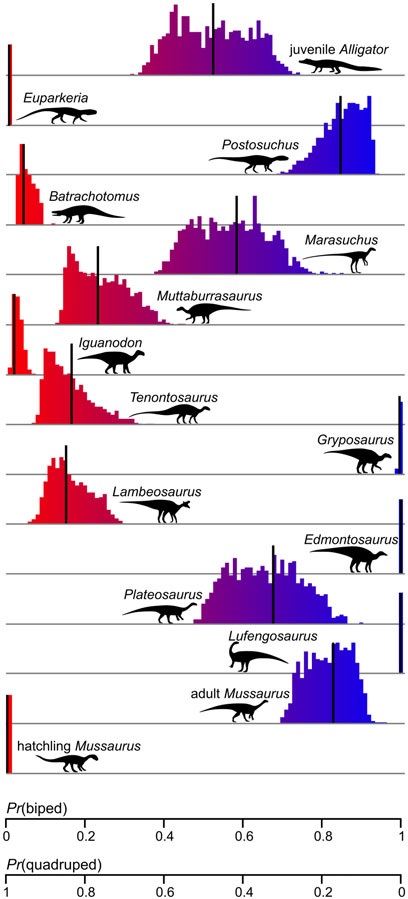
We also had some non-Triassic/Jurassic species in our sample that produced interesting results. Broadly, we obtained good support for the conclusion that “duckbill” ornithopod dinosaurs (Muttaburrasaurus, Tenontosaurus, Iguanodon, Lambeosaurus) were usually quadrupedal, as their hooved “hands” and fossil footprints strongly indicate; but two species (Gryposaurus and Edmontosaurus) ended up (mis)classified as bipeds; a result that analogous methods based on bone dimensions also tend to get. Hence we suspect that simply relying on parameters such as these studies’ to estimate locomotor posture is insufficient. Ornithopods had unusually short, skinny forelimbs that seem to mislead such analyses.
Intriguingly, we found more variation in body shape overall in bipeds vs. quadrupeds (Video 1), and a trend for linked forelimb length and centre of mass position (i.e. more front-heavy) in bipeds; but that bipeds tend to have longer hindlimbs and a centre of mass closer to the hips than quadrupeds do. This makes sense in terms of balance: bipeds need to stand and move with their centre of mass over their two feet, and longer legs help them both balance and move quickly. The origin of flight in birds helped “decouple” the influence of body dimensions on bipedalism, via a change in limb posture to more crouched, as our data supported.
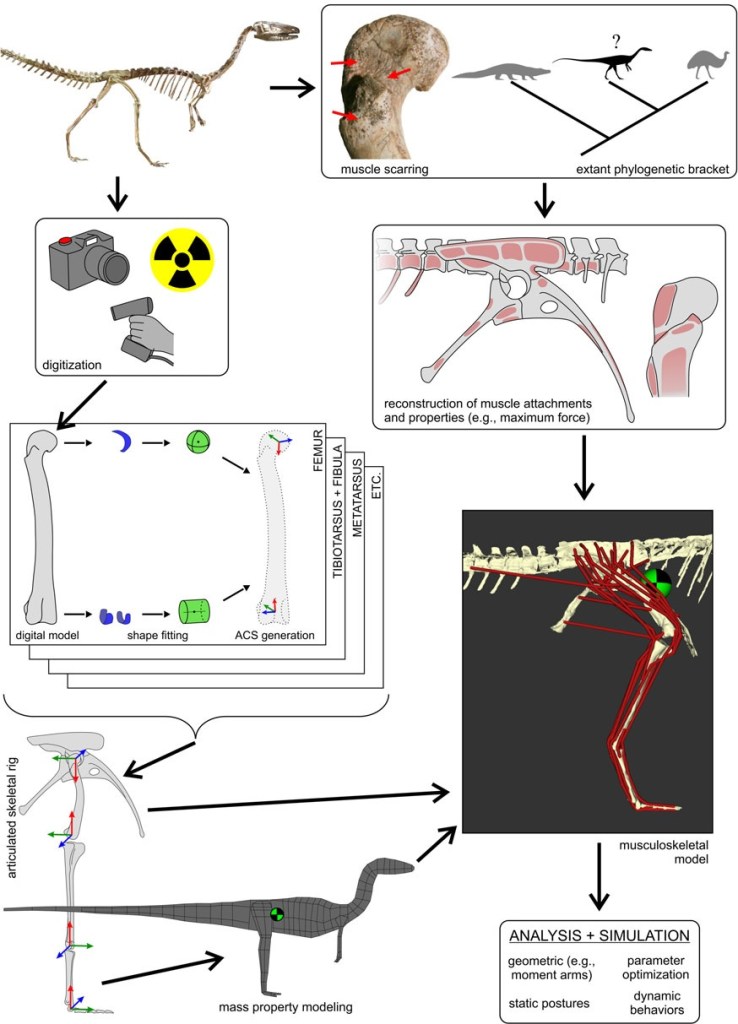
The second paper laid our a new refined method for building 3D computer models of whole archosaurs, including reconstructed muscles, and used this method to estimate how fast the bipedal Triassic dinosaur Coelophysis might have run. We explained how we go from 3D scanned bones to an articulated, skeletal “digital puppet”, providing new tools to do this more objectively by semi-automatically defining joint surfaces using the bony geometry. Next, we provided an outline of how one “fleshes out” the skeleton, adding masses and centres of mass and inertia, as well as then providing muscular attachments and connecting those to create a full-fledged 3D musculoskeletal model. Such a model can then be given physiological properties based on its anatomy and data from living animals, for which we gave new methods to do so.
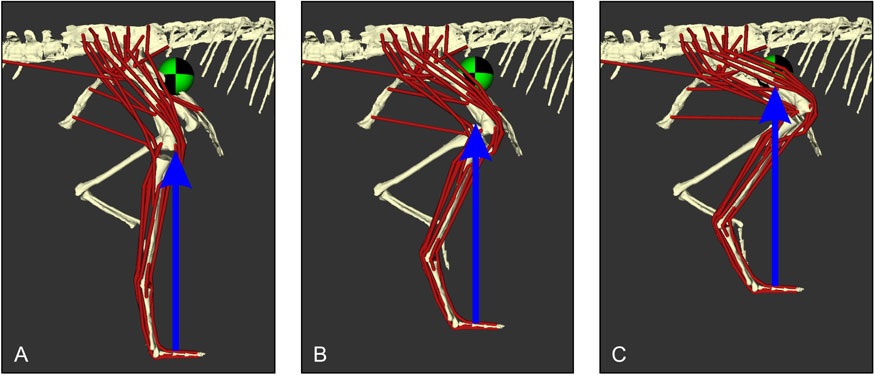
Finally, we used open-source, well-vetted biomechanics software OpenSim to simulate what speeds the hindlimb musculature could have sustained. First, we found that applying this method to good models of a human and ostrich in sprinting poses gets the “correct” conclusion that they could indeed run that way, although with the interesting finding that their knee muscle strength, not ankle muscles as our prior studies had inferred, might be the major limit on speed.

Second, for the 13kg Coelophysis, our range of models (Figure 6) indicated that it could have run fairly quickly if it used relatively straight legs; independently supported by fossil footprints and other prior predictions from its anatomy. Hence our analyses of sprinting in ostriches, humans and Coelophysis gave good confidence that we can reconstruct running ability in small- to moderate-sized bipeds with our novel methods.
These two new papers move our DAWNDINOS research into an important new stage where we will begin testing hypotheses about archosaurian locomotor evolution and its links with the Triassic-Jurassic transition.
For another take on the second paper, see the journal’s Featured Article story on it!
